Cart
Cart

Serotonin
26 Dec 2016
Serotonin is the biogenic amine which is formed of tryptophan amino acid by its hydroxylation and decarboxylation. A significant amount of a serotonin contains in the enterokhromaffinnykh cells of intestine, CNS, mainly in hypothalamus and mesencephalon, thrombocytes, smaller quantity in the labrotsitakh, mast cells, adrenals. A serotonin exerts impact on nervous activity, causes reduction of a smooth musculation of an intestine, uterus, bronchi, and also narrowing of vessels. Reactions of an organism to a serotonin are the cornerstone the central, myotropic, ganglionic, reflex effects.
Education. A serotonin (5-gidroksi-triptamin, 5-HT) is synthesized in the enterokhromaffinnykh intestine epithelium cells from a L-tryptophan. A serotonin is formed also in nervous cells of mezenterialny plexus and in a CNS where plays a mediator role. Thrombocytes don't synthesize a serotonin, however take it and accumulate.
Serotonin receptors. There are several types of receptors. From the pharmacological point of view receptors 5-HT1 and 5-NT2, and also 5-HT3 and 5-HT4 are important. As a rule, receptors work through G-protein. The receptor 5-HT3 represents cationic channel.
Effect of serotonin. Cardiovascular system. Influence of serotonin on cardiovascular system is complex as serotonin has effect (sometimes opposite referred) in many sites through different types of receptors. Through 5-HT2-retseptory located in unstriated muscles of walls of vessels, a serotonin causes narrowing of vessels. At the same time indirect effect of serotonin can lead to vasodilatation and depression of the ABP: through 5-HT1A-retseptory it blocks sympathetic neurones of a brainstem and the periphery that leads to depression of a sympathetic tonus; activation of 5-HT1 receptors in a vascular endothelium leads to emission of vazodilatator (prostatsickin, NO). The serotonin which is released from thrombocytes plays a role in processes of thrombogenesis, hemostasis and in hypertonia pathogenesis at pregnant women.
Sumatriptan (an agent against migraine) is an agonist 5-HT1D and 5-HT1B-retseptorov. The narrowing of cranial vessels caused by blockade of emission of neuropeptids leads to a so-called "neurogenic inflammation" or to direct vascular crisis. At the same time there can be a feeling of constraint in a breast bound to a spastic stricture of coronary arteries. Other "tryptones" are naratriptan, zolmitriptan and rizatripan.
Digestive tract. The serotonin formed in mezenterialny plexus or enterokhromaffinnykh cells stimulates a peristalsis and secretion of an intestine by means of influence on 5-HT4-retseptory.
Attempts of pharmacological impact on a motility of an intestine through serotonin receptors were ineffective so far. Tsizaprid, an agonist 5-HT4, however, can increase propulsive motility of an intestine. This side effect is blocked by Atropinum that demonstrates rising of concentration of Acetylcholinum under the influence of tsizaprid. Drug renders a set of side effects. It is inactivated on the mechanism with participation of SURZA4 that leads to various unforeseen interactions with other substances. Tsizaprid causes disturbances (including serious) a cordial rhythm (reduction of QT intervals) therefore drug didn't find application.
Central nervous system. Serotonin receptors play an important role in the CNS various functions.
Fluoxetine is the antidepressant blocking the return serotonin reuptake. It considerably increases motivation, and also lowers appetite.
Sibutramin, blocker of the return neuronal serotonin reuptake and Noradrenalinum; it is applied as remedy for obesity.
Ondansetron has the expressed antiemetic effect at the vomiting caused by application of tsitostatik. He is an antagonist of 5-HT3-retseptorov. Analogs of ondansetron are tropisetron and granisetron.
Acid and others of psychedelia (psikhotomimetika), such as mescaline and psilocybin, cause hallucinations, disorders of consciousness, fear, perhaps, because of activation of 5-NT-retseptorov.
Effects of serotonin
Staff of University of McMaster became authors of scientific work during which the key gene of obesity - a kind of serotonin has been found. It is well-known that serotonin is "joy hormone", his development by a brain promotes emotional stability and good mood. But, as the Canadian researchers explain, the serotonin which is responsible for pleasant emotions belongs to the first type of this connection.
"He is divided into two types: in the scene of action and in a synthesis form. The first type is developed in a brain and influences different emotions", - biologists have explained.
To the second type, peripheral serotonin belongs this substance regulates activity of brown fatty tissue on which development of obesity depends. You can also like Pankramin.
Brown fat contains components which promote decrease in sugar in blood, and also to burning of calories and their processing in energy. On a body of the person there are certain zones where brown fat settles down and his cages are more active, the figure at the person is more harmonious. The Canadian researchers have come to conclusion that suppressing serotonin of the second type; it is possible to increase considerably metabolic activity of cells of brown fatty tissue. She, in turn, will force a body "to burn" white fat � and, it will occur irrespective of how high-calorie food is eaten by the person.
Serotonin receptors
Effects of serotonin are extremely various. This substance serves as a mediator in TsNS, influences smooth muscles of vessels and a GIT, participates in vascular trombotsitarnom a homeostasis. Methods of molecular cloning revealed unexpectedly large number the serotonin receptors which based on structure and function can be divided into 4 types. 5-HT1-, 5-HT2-and 5-HT4-retseptory are integrated to G-proteins and through these proteins and the corresponding systems of the second intermediaries influence functions of various enzymes and elek-trofiziologich properties of effector cages. On the contrary, 5-HT3-retseptory are tied with ion channels. Here we will consider stimulators and blockers the serotonin receptors. The newest medicines of these groups which are selectively operating on separate subtypes the serotonin receptors were received in works with use of recombinant receptors. We will stop also on experimental models which apply to a research of the means influencing difficult mental functions and their violations persistence, aggressive behavior, uneasiness, depression, cycle dream wakefulness and other. Modern selective stimulators of separate subtypes the serotonin receptors already with success are applied in case of migraine and uneasiness, and the selective blockers in case of a number of gastrointestinal violations. It is possible to influence physiological effects of serotonin also by means of the means operating on serotoninergich transfer. So, inhibitors of the return capture of serotonin were effective medicines for treatment of depression and uneasiness.
In spite of the fact that the serotonin role in many physiological and pathological processes doesn't raise doubts, points of its application and mechanisms of action are studied badly. Perhaps, such situation is partly caused by variety the serotoninovykh of receptors. These receptors taped in the beginning by pharmacological methods are received by KDNK cloning today. Recombinant serotoninovy receptors use for studying of molecular mechanisms of effect of serotonin, and also for search of the agents which are selectively influencing separate subtypes of these receptors. The circle of a clinical use of similar agents becomes wider and wider.
Historical information
In the 1930th Erspamer began to study localization of enterokhromaffin cells by means of stains on indole derivatives. The highest concentration of such derivatives was taped in mucous by a GIT; further there were thrombocytes and some departments of a CNS (Erspamer, 1966). After a while Peydzh and coauthors, working in the Clevelend clinic, for the first time emitted the vasoconstrictive substance released from thrombocytes in the course of a bleeding stopping and deciphered its structure (Rapport et al., 1948). This substance named by Paige a serotonin (Page, 1976), appeared to those derivatives of an indole which was investigated by Erspamer. The description of ways of synthesis and disintegration of a serotonin (Uden-friend, 1959) and its angiotonic properties (Sjoerdsma, 1959) allowed to make a hypothesis according to which implications of a so-called carcinoid syndrome at patients with tumors from enterokhromaffin cells are caused by the increased production of this substance. Really, such patients have a daily ejection with urine of a serotonin and its metabolites can reach hundreds of milligrams. Some symptoms of this disease to some extent indicate mechanisms of effect of serotonin. So, at patients the psychosis similar to the acid arising at reception can develop. Considering that in animal and vegetable tissues substances with hallucinogenic action are found similar with tripta-miny, it is possible to assume that similar substances are formed and cause a psychotic symptomatology in patients with characinoid syndrome. About mediator function of a serotonin in a brain of mammals it was suggested in the mid-fifties (Brodie and Shore, 1957).
The first data on molecular mechanisms of effect of serotonin were obtained in experiences on a liver of Fasciola hepatica (Mansour, 1979). Under the influence of serotonin at it mobility and concentration of tsAMF sharply increased; both that, and another effects were blocked by an acid. Increase in mobility was caused by tsAMF-dependent phosphorylation of fosfofruktokinaza limiting glycolysis enzyme. However the serotoninovy receptors mediating these effects at a liver, probably, others, than receptors of mammals interfaced with adenilattsiklaz to serotonin. At the latest so detailed data on mechanisms of effect of serotonin it wasn't succeeded to receive so far.
Serotonin appeared at plants and animals already at early stages of evolution, and to these, perhaps, the abundance of serotonin receptors speaks (Peroutka and Howell, 1994). The cloning of these receptors showed that some medicines which were earlier considered selective in relation to their separate subtypes actually have high affinity in relation to several subtypes. In more detail about history of studying and effects of serotonin see the article Sjoerdsma and Palfreyman (1990).
Chemical properties of serotonin
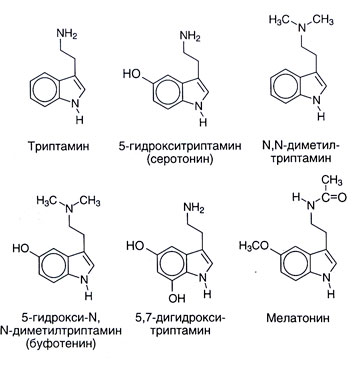
Sources. The chemical structure of serotonin and some close connections to it is given in fig. 11.1. Serotonin is widespread in a plant and animal life: it is found at vertebrata, tunicates, mollusks, arthropods, coelenterates, in fruit and in nuts. It is found also in poisons � in a nettle, in wasps and scorpions. Numerous synthetic or natural relatives to substance serotonin also in a varying degree have the central and peripheral physiological effects. Many N-or O-metilirovannye indolamines (for example, N, N-dimetiltriptamin) are hallucinogens. As they can be developed in an organism, long considered them possible responsible for at least some displays of psychoses. Melatonin (5-metoksi-N-atsetiltriptamin) is formed of serotonin by N-acetylation with the subsequent O-methylation (fig. 11.2). This substance serves as the main indolamine of body where its synthesis is regulated by external factors (in particular, illumination level). Melatonin causes depigmentation of melanotsit of skin and suppresses function of ovaries. Perhaps, it plays a part in biorhythms and therefore it can be useful at a syndrome of change of time zones.
Synthesis and catabolism. A serotonin is formed of irreplaceable amino acid of a tryptophan in 2 stages (fig. 11.2). At the first stage under the influence of a triptofangidroksilaza 5 hydroxytryptophan are formed, it is the limiting serotonin synthesis reaction. Triptofangidroksilaza represents an oxidase with the admixed functions. Molecular oxygen, and as coenzyme tetrahydrobiopterine takes part in the reaction catalyzed by it. Activity of triptofangidroksilaza, as well as tyrosinehydroxylase, is regulated by phosphorylation, however the triptofangidroksilaza isn't inhibited by the final product on the mechanism of negative feedback. In a brain of triptofangidroksilaz it isn't saturated with substrate and therefore rate of synthesis of a serotonin depends on concentration of a tryptophan. The last comes to brain cells by active capture by means of the carrier which is responsible for transport of several neutral and branched amino acids. In this regard the maintenance of a tryptophan in a brain depends not only on its concentration in plasma, but also on concentration of other amino acids competing with a tryptophan for a carrier.
Decarboxylation leads 5 hydroxytryptophans to formation of a serotonin. The long dispute on whether decarboxylases 5 hydroxytryptophans and DOFA are different or the same enzyme, was resolved by KDNK cloning methods it turned out that the same gene product is responsible for decarboxylation of both substrates. Now this enzyme is called decarboxylase of aromatic L-amino acids. It is extremely widespread and affects many substrates. 5 hydroxytryptophan are decarboxylized very quickly and in a brain it is almost not found. In this regard attempts to affect concentration in a serotonin brain by concentration change 5 hydroxytryptophans are doomed to failure.
The main way of catabolism serotonin the transformation into 5-hydroxyindolacetic acid which is also proceeding in 2 stages (fig. 11.2). At first under the influence of MAO the 5-hydro-xyindolacetaldehyde which then passes into 5-gidroksi-indolacetic acid under the influence of widespread aldegiddegidrogenaza enzyme in an organism is formed (the insignificant quantity 5 hydroxyindolacetaldehydes turns into alcohol � 5-gidroksitriptofol). 5-hydroxyindolacetic acid is activly removed from a brain; this process is suppressed with a nonspecific inhibitor of transepithelial transfer probenetsidy. As in nervous cells nearly 100% of all metabolites of a serotonin fall to the share of 5-hydroxyindolacetic acid, rate of a circuit of a serotonin in a brain is estimated on rising of level of 5-hydroxyindolacetic acid after introduction of probenetsid. 5-hydroxyindolacetic acid which is formed in a brain and other organs, and also small amounts of a 5-gidroksitriptofol and glucuronides are removed with urine. Normal the daily egestion of 5-hydroxyindolacetic acid at the adult makes 2 � 10 mg. Higher values � a reliable sign of carcinoid syndrome. Sharply increased synthesis of a serotonin at this disease demands large numbers of pyridinic nucleotides and a tryptophan and therefore signs of deficiency of nicotinic acid and a tryptophan not a rarity at such patients. Ethanol causes rising of maintenance of NADN, and as a result of 5 hydroxyindolacetaldehyde passes from an oxidizing way of a catabolism to recovery (fig. 11.2).
These are raises egestion of a 5-gidroksitriptofol a little and respectively reduces egestion of 5-hydroxyindolacetic acid.
There are two isoenzymes of MAO: MAO A and MAO B. At first they were divided based on affinity to substrata and sensitivity to inhibitors; now both isoenzymes are cloned, and properties of the cloned and natural forms were identical (Shih, 1991; see also hl. 10). MAO A has preferential affinity to serotonin and noradrenaline, and its selective inhibitor is clorgilin. MAO B more works on β-fenil-etilamin and benzylamine; the MAO B selective inhibitor selegilin. Affinity of both isoenzymes to dopamine and triptamin is identical. Nervous cages contain both MAO A, and MAO B � mainly on an external membrane of mitochondrions. As the main isoenzyme of platelets which also contain serotonin in high concentration serves MAO B.
It was supposed that there are also other ways of a catabolism of serotonin, for example sulphation and About - or N-methylation. The final journey, in particular, could lead to formation of an endogenous psychotropic substance 5-hydroxy-N, N-dimetiltriptamina (bufotenina, fig. 11.1). However other metilirovanny indolamines (N, N-dimetiltriptamin, 5-metoksi-N, N-dimetiltriptamin) have much more expressed hallucinogenic properties, and their role in pathogenesis of psychoses is more probable.
The inactivation of serotonin is performed not only by enzymatic disintegration, but also by means of the return capture. Na + - the dependent carrier located on an external surface of presynaptic membrane of the serotoninergich termination (provides removal of serotonin from a synoptic crack) and an external surface of a membrane of platelets is responsible for this capture (extracts serotonin from blood). At platelets it is the unique way of replenishment action of serotonin as there are no enzymes of synthesis of this substance in them. The serotonin carrier, as well as other carriers of monoamines, is cloned (hl. 12).
Points of serotonin application
The most part of all maintenance of serotonin in an organism falls to the share of peripheric tissues though it serves also as a mediator in a CNS. Its concentration in enterokhromaffinnykh cells and thrombocytes is highest. A serotonin plays an important role in a regulation of a motility of a GIT.
Enterokhromaffinny cells. These cells settle down in mucous a GIT. Especially there is a lot of them in a duodenum. In the enterokhromaffinnykh cells it is synthesized from a tryptophan and a serotonin collects, and also other biologically active agents, for example substance P and kinina contain. There is a certain level of basal secretion of a serotonin in a GIT. This secretion amplifies at mechanical stretching (for example, when entering a nutrition or hypertonic salt solution) and at a boring of motive fibers of vagus nerves. Perhaps, the stimulating effect of serotonin on a motility of a GIT is mediated also by its influence on neurones of an intermuscular plexus (Gershon, 1991; see also hl. 38). Sharply increased secretion of a serotonin and other biologically active agents at carcinoid syndrome is followed by the corresponding gastrointestinal, cardiovascular and nervous disturbances. Besides, the increased synthesis of a serotonin can result in deficiency of nicotinic acid and a tryptophan.
Thrombocytes. Thrombocytes differ from other formulated elements of a blood, in particular, in ability to take store and release a serotonin. Synthesis of a serotonin in thrombocytes doesn't happen. A serotonin is taken thrombocytes from a blood and comes for storage to secret elektronop granules by means of active transport. These processes are in many respects similar to capture and storage of Noradrenalinum in the sympathetic terminations (hl. 6 and 12). Through a membrane of thrombocytes a serotonin is transferred by means of Na + - dependent transport, and to granules � by secondary active transport with use as an energy source of an electrochemical gradient for H+ framed by N +-Atfazoy. At the same time concentration of a serotonin in granules reaches 0.6 mol/l, it is 1000 times higher, than in cytoplasm of thrombocytes. Rate On + - dependent serotonin reuptake by thrombocytes a sensitive indicator of activity of inhibitors of serotonin reuptake.
The main function of thrombocytes is hemostasis: they close gaps in the damaged endothelium. On the other hand, the integrity of an endothelium plays an important role in functioning of thrombocytes (Furchgott and Vanhoutte, 1989). The endothelium constantly contacts to thrombocytes as because of the shift forces operating in the current blood they are displaced to the periphery of vessels (Gibbons and Dzau, 1994). Vasoconstrictive effect of serotonin and tromboksan of A2 is resisted by an endothelial factor of vasorelaxation (N0 and, perhaps, some other substances) (Furchgott and Vanhoutte, 1989; fig. 11.4). For adhesion and aggregation of thrombocytes the condition of an endothelium has crucial importance (Hawiger, 1992; Ware and Heistad, L993). When thrombocytes adjoin to the damaged endothelium, they emit the substances causing their adhesion and release of a serotonin. ADF and ���������� A2 belong to such substances (hl. 26 and 55). Linkng of a serotonin with 5-HT2A-retseptorami has the weak proagregant effect which is sharply amplifying in the presence of a collagen. If defect of a vascular wall reaches smooth muscle layers, then a serotonin renders the direct vasoconstrictive effect serving as one of hemostasis mechanisms. This effect amplifies under the influence of biologically active agents which are emitted in the field of damage tromboksan of A2, kinin, vasoactive peptides. Formation of thrombi at an atherosclerosis is promoted by destruction of an endothelium and, as a result, lack of an endothelial factor of vasorelaxation. In these conditions the processes conducting to thrombogenesis proceed uncontrolledly, as a vicious circle. A part in them is played also by a serotonin. The similar picture can be observed at other illnesses of vessels, for example Reynaud's syndrome and vasospastic stenocardia.
The description to fig. 11.4. Functions of a serotonin of thrombocytes. Release of a serotonin from thrombocytes is started them by adhesion and aggregation. In turn, a serotonin causes 1) activation �-��-receptors thrombocytes and, vrezultata, change of a form and acceleration of aggregation of the last, 2) activation 5-HT, - similar receptors of an endothelium with allocation of an endothelial factor of vasorelaxation, 3) activation of S-HT-receptors of unstriated muscles of vessels and narrowing of the last. All these processes proceed in interaction with many other biologically active agents and eventually lead to bleeding stopping.
Cardiovascular system. Typical reaction of blood vessels to serotonin is narrowing. Vessels of organs of GIT, kidneys, lungs and brain are especially sensitive to it. Serotonin causes also reduction of unstriated muscles of bronchi. Its effects on heart are various that serotonin receptors change of tonus of vegetative nerves and reflex reactions is explained by activation of different subtypes (Saxena and Villalon, 1990). So, direct positive chronotropic and inotropic effect of serotonin on heart can be disguised by effects of exaltation of the fibers going from bar receptors and chemoceptors. Influence of a serotonin on the afferent terminations of vagus nerves causes Betsold's reflex Yarisha who is shown a sharp bradycardia and falling of the ABP. Sometimes arterioles under the influence of a serotonin aren't narrowed, and, on the contrary, suppression of release of Noradrenalinum from the sympathetic terminations extend as a result of allocation of an endothelial factor of vasorelaxation and Prostaglandinums, and also. On the other hand, a serotonin in itself strengthens vasoconstrictive action of Noradrenalinum, angiotensin 11 and Histaminum. It promotes even more effective haemo static effect of serotonin (Gershon, 1991).
GIT. Probably, as the main source and storage of a serotonin in an organism serve enterokhromaffinny cells mucous a GIT. The serotonin allocated by these cells comes through a portal vein to a liver where it is metabolized under the influence of MAO A (Gillis, 1985). Some amount of serotonin passes a hepatic metabolism, but is quickly taken an endothelium of pulmonary capillaries and also is affected by MAO. The serotonin which is allocated in a wall of organs of a GIT at their mechanical stretching or exaltation of vagus nerves participates in a local regulation of these organs. Under the influence of a serotonin a motility of a stomach and an intestine can both amplify, and to be braked (Dhasmana et al., 1993) as in a GIT there are at least 6 subtypes the serotonin receptors (tab. 11.2). Stimulating effect of serotonin is caused by its action on the terminations of the nerves suitable to longitudinal and circular muscular layers (5-HT4-retseptory), on intramural neurons (5-HTj-and 5-NT|R-retseptory) and is direct on smooth muscles (5-��-receptors in intestines and 5-HT2B-retseptory in day of a stomach). In a gullet serotonin affects 5-HT4-retseptory that at different types of animals can be followed both by reducing, and relaxation of smooth muscles. 5-HT3-retseptory (it is a lot of attendees at the terminations of sensitive fibers of the wandering and other nerves, and also on the cages) play a key role in an emetic reflex (Grunberg and Hesketh, 1993). In an intermuscular texture the serotoninergetic terminations are found. Release of serotonin in intestines is caused by acetylcholine, irritation of sympathetic nerves, and increase in intra intestinal pressure and decrease in pH (Gershon, 1991). The serotonin which is emitted at the same time, in turn, starts peristaltic reducing.
The main area of concentration of serotonin neurons bodies in CNS are cores of seam of brainstem. Processes of these neurons go to all departments of a head and spinal cord (hl. 12). A serotonin is allocated not only in the presynaptic terminations, but also in a so-called varicosity of axons where there are no accurately expressed synapses (Descarries et al., 1990). In these cases it affects many adjacent structures at once. Such feature of allocation and effect of serotonin will be compounded with the widespread point of view that a serotonin is not only a mediator, but also the neuron modulator (hl. 12).
In the terminations of serotonin neurons there are all components necessary for synthesis of a serotonin from a tryptophan (fig. 11.2). The formed serotonin quickly comes to synoptic blisters where on it MAO can't work. After release in a synoptic cleft a serotonin is again taken the nervous termination by means of Na - a dependent carrier. This return capture serves as an effective way of an inactivation of a mediator. The same molecules of a serotonin which don't come back to the nervous termination are blasted by MAO located in postsynaptic neurons and the next cells.
Electrophysiological effects. These effects of a serotonin differ in different areas of a brain and in different neurons, and depend on what receptors it affects (tab. 11.3; Aghajanian, 1995). A serotonin can have the exciting and brake effect differing on temporary dynamics on the same neurons. So, in hippocampus neurons a serotonin causes at first the hyper polarization (caused by activation of 5-HT 1A-receptors), then the slow depolarization (caused by activation of 5-HT4-retseptorov).
TsNS. Serotonin influences the TsNS many functions, including a dream, cognitive activity, perception, management of movements, thermal control, painful sensitivity, appetite, sexual behavior and endocrine regulation. In a brain all cloned serotonin receptors are found, and often at the same department there are several such receptors. Moreover, though the expression of serotonin receptors in separate neurons is studied insufficiently, it is possible to believe that on the same neuron several subtypes of these receptors can be located, and their activation can be followed by both synergistic and antagonistic effects. It can be the cause of an extraordinary variety of influences of serotonin on brain functions.
Hyper polarization and decrease in resistance of a membrane arising in case of activation 5-HT) A-receptors, are caused by increase in potassium permeability. These effects are blocked by toxin, but don't depend on tsAMF. Therefore, they can be caused by direct interface of 5-NT|A-retseptorov (through the G-protein similar to Gi) with the potassium channel (Andrade et al., 1986). Activation of the receptors located on a body and dendrites of neurons of kernels of a seam of a trunk of a brain also leads to K+ - dependent hyper polarization. Here too participates in a broadcast j of a signal from a receptor to the channel sensitive to G-protein toxin, but potassium current has other characteristics, than that which arises in case of activation of postsynaptic 5-HT1A-retseptorov in a hippocampus. It isn't known yet by means of what mechanisms activation 5-HT1D-ayTopeuerrropoB leads to serotonin release suppression. Probably, the number of the calcic channels opening in response to nervous impulse the potential decreases.
Activation of 5-HT2A-retseptorov is followed by slow depolarization. In some departments of a brain (for example, in prefrontal cortex, an adjacent kernel and a motive kernel of a facial nerve) this depolarization is caused by decrease in potassium permeability (Aghajanian et al., 1987). There is also other mechanism connected with influence on ion channels; it leads to growth of excitability of neuron and strengthening of reaction to exciting mediators (for example, a glutamate). The role of fosfoinozitid system in these effect isn't established yet. When on the same neuron there are also 5-HT1-retseptory and 5-Nt2d-retseptory, final reaction to serotonin depends on a ratio between the hyper polarization caused by activation of 5-HT1-retseptorov, and the depolarization caused by activation of 5-HT2A-retseptorov. Against the background of 5-Nt2d-blokatorov hyper polarization amplifies. In many areas of bark 5-HT2A-retseptory are located on Gamkergichesky inserted neurons and on pyramidal neurons.Therefore, stimulation of these receptors can lead to multidirectional influences on pyramidal neurons in dependence because whether action on these neurons or on Gamkergichesky neurons will prevail. Activation of 5-Nt2s-retseptorov on oocytes of frog, the express of m of RNA of these receptors, leads to suppression of potassium current. In a brain such effect isn't revealed yet. The stimulation of 5-HT4-retseptorov which is followed by activation also causes the slow depolarization caused by decrease in potassium permeability in neurons. It isn't clear yet why two different types of serotonin receptors interfaced to various systems of the second intermediaries render the same physiological effect. Moreover, slow depolarization is caused by activation of one more type of serotonin receptors 5 HT1P-receptors. These receptors are accompanied by available only on intra intestinal neurons and have special pharmacological properties (Gershon, 1991).
Activation of 5-HT3-retseptorov causes bystry depolarization. It is caused by opening of the channel passing Na + and K+ (Higashi and Nishi, 1982) and making a uniform complex (hemo sensitive channel) with a receptor. The similar organization of a 5-HT3-retseptora similar to the N-holinoretseptora organization has been confirmed in experiences with local fixing. 5-HT3-retseptory are found in TsNS, sympathetic ganglia, parasympathetic and sympathetic afferent fibers, intra intestinal neurons and cellular lines of neyronal origin (for example, NG108-15). On the pharmacological properties 5-HT3-retseptory differ from others serotonin receptors; perhaps, there are several subtypes of these receptors differing in various combinations of subjunits.
Mental functions. The means operating on serotonin receptors and various changes of mental functions. Many experimental models intended for a provisional estimate of the stimulating or blocking activity of medicines in relation to these or those receptors, are based on a research of such stereotypic motive acts as, for example, reflex. Behaviouristic techniques (for example, a medicine choice method) allow to suggest about a subjective component of reactions to psychotropic drugs. These techniques are used also for a research of the medicines operating on serotonin transfer, in particular hallucinogens (see below). The analysis of enormous number of the works devoted to influence of serotonin on behavior is beyond our book, and we will stop only on those experiments which have a direct bearing on psychopathology of the person. For more detailed acquaintance with this subject it is possible to recommend the fine reviews Glennon et Lucki (1988), Zifa and Fillion (1992), Koeketal. (1992).
Cycle a dream is wakefulness. Regulation of a cycle a dream is wakefulness became one of the first mental functions for which the serotonin role was accurately established. After the classical work on cats which is carried out by Mouret and ����. (Mouret et al., 1967), the set of data that the serotonin stock depletion by means of causes the sleeplessness eliminated by entering of a predecessor of serotonin 5 of appeared. It appeared also that tryptophan and not selective stimulators of serotonin receptors shorten time of falling asleep and extend the general duration of a dream. Blockers of serotonin receptors can both rise, and to lower a share of a deep slow sleep that, probably, is caused by action on different subtypes of serotonin receptors (Wasquier and Dugovic, 1990). Both at animals, and blockers 5-HT2A-and 5-Nt2s-retseptorov (for example, riganserin) rather reliably cause increase in a share of a deep slow sleep.
Aggression and impulsiveness. The data obtained both on animals and on the person, demonstrate that a serotonin plays an important role in aggressive and impulsive behavior. In many clinical trials communication between the low level of 5-of hydroxyindolacetic acid in SMZh and such behavior is shown (Brown and Linnoila, 1990). So, depression of this level is bound to impulsive attempts of suicide (but not with suicidal thoughts; Virkkunen et al., 1995). As well as in case of all other effects of a serotonin, animals have no final data on influence of a serotonin on aggressive behavior yet though the assumption of such influence is quite proved. Recently there were genetic data supporting and dilating such views. Serotonin receptors investigated by methods of genetic engineering 5-HT) the V-receptor was the first of. By a homologous recombination the line of mice who have a gene coding this receptor was received it was inactivated (Saudau et al., 1994). At such animals the sharpest aggression developed that speaks about a role of these receptors or in a becoming of the neuronic contours which are responsible for aggressive behavior or it is immediate in the most such behavior. At the person the dot mutation of the gene coding MAO A is taped; at the same time extreme aggression in combination with mental retardation is also observed (Brunner etal., 1993). As it appeared, the corresponding implications are available also for mice with the mutations resulting in deficiency of MAO A (Cases et al., 1995). These data, certainly, confirm a hypothesis of a role of disturbances of serotoni-nergic transfer in aggressive behavior.
Uneasiness and depression. Efficiency of the means influencing serotoninergichesky transfer (for example, inhibitors of the return capture of serotonin), in case of disturbing frustration and depression the evidence for benefit of a serotonin role in pathogenesis of these conditions. However on classical experimental models of these frustration receive ambiguous results the effect of medicine depends on species and breed of an animal and the applied technique. So, partial agonist of 5-HT1A-retseptorov buspiron (hl. 19), being an effective tranquilizer at the person, doesn't reduce at animals uneasiness in the experiences delivered by a technique of preference or avoiding; meanwhile this technique was used in case of development of tranquilizers. At the same time buspiron and other blockers of 5-HT1A-retseptorov have quite effective anxiolytic effect on other models of disturbing frustration (Barrett and Vanover, 1993). Recent works on mice with the inactivated genes of 5-NT1L-retseptorov also witness for benefit of a role of these receptors in pathogenesis of uneasiness and, perhaps, depressions (Parks et al., 1998; Ramboz et al., 1998). On the other hand, stimulators of some the serotonin receptors, including 5-HT2A-, 5-HT2C-and 5-HT3-retseptorov cause uneasiness both in experimental animals, and in the person. The role of these receptors and in experimental models of a depression is supposed (for example, the trained helplessness).
The person of direct data on a serotonin role in a pathogenesis of a depression has still not enough. At the same time there are very bright clinical facts. So, at patients with a depression effects of inhibitors of the return serotonin reuptake quickly are eliminated at the influences reducing serotonin level in a brain. As such influences serve, for example, reception of parachlorphenylalanine or the drinks which aren't containing a tryptophan, but rich with neutral amino acids (Delgado etal., 1990). It is interesting that these influences in itself don't cause and don't aggravate a depression. It means that the sufficient level of a serotonin in a brain is necessary first of all for efficiency of inhibitors of its return capture.
Agents influencing serotonin level in tissues
In the experiments referred on clarification of a physiological role of a serotonin it is possible to apply either blockers of serotonin receptors, or the agents influencing serotonin level in tissues. Until recently in the majority of works only drugs of the second of the mentioned groups were used mechanisms of action of blockers the serotonin receptors were studied badly.
At the low maintenance of a tryptophan in a diet serotonin level in a brain decreases, at the high content of tryptophan increases. As the limiting reaction of synthesis of a serotonin is catalyzed, inhibitors of this enzyme cause sharp falling of level of a serotonin. Most often use an irreversible selective inhibitor of a triptofangidroksilaza parachlorphenylalanine. Under the influence of this substance serotonin level considerably and for a long time decreases, and the maintenance of catecholamins doesn't change.
Parachloramphetamine and other halogenated amphetamines cause emission of serotonin from thrombocytes and neurons. In a brain after that there comes long depression of reserves of serotonin. Halogenated amphetamines are widely used in experimental works. Two of them fenfluramin and decsfenfluramin were applied as anorecsant, however in connection with reports on their cardio toxic action in 1998 they in the USA were withdrawn from sale. Consequences of use of these agents are up to the end not studied. In the serotonin brain neurons they cause expressed and long (up to several weeks) depression of level of a serotonin, and the content of proteins, specific to these neurons a carrier of a serotonin and triptofangidroksilaza at the same time decreases. It can demonstrate neurotoxin action, however signs of death of neurons under the influence of halogenated amphetamines aren't found. Derivatives of triptamin with additional deputies in Indo-flax a ring (for example, 5.7-digidroksitriptamin; see fig. 11.1) undoubtedly cause death the serotonin neurons. Introduction of a 5.7-digidroksitriptamin an adult animal is led to selective destruction by serotonin terminations, however bodies of neurons don't suffer, and over time the terminations regenerate. On the contrary, at newborn animals both the terminations, and bodies of serotonin neurones and therefore neogenesis doesn't happen perish.
Inhibitors of the return serotonin reuptake, for example fluoxetine belong to the agents which are precisely influencing serotonin transfer. The mechanism of their action consists in extension of effects of the serotonin allocated at initiation of the nervous terminations. If along with these drugs to enter 5 hydroxytryptophan, then serotonin influences sharply amplify. Inhibitors of the return serotonin reuptake are one of the most modern and widespread antidepressants. The inhibitor of the return serotonin reuptake, Noradrenalinum and Dofaminum sibutramin is applied as anoreksant. In an organism two active metabolites which, probably, and have therapeutic effect are formed of it. It isn't clear yet, influence on what mediator caused effect of sibutramin.
Inhibitors of MAO and Reserpinum belong to not selective agents influencing serotonin level in tissues. Inhibitors of MAO block the main way of a metabolism of a serotonin, and Reserpinum causes its emission from neuron depot with the subsequent attrition. All these agents lead to the expressed depression of maintenance of serotonin, however at the same time in the same degree also the level of catecholamine decreases. Therefore as agents for the pharmacological analysis inhibitors of MAO and Reserpinum are used seldom. They were applied in psychiatry: Reserpinum as narcoleptic, and MAO inhibitors as antidepressants.